Journal Publications
|
Yang, Y., Yared, D.G., Fortune, E.S. and N.J. Cowan (2024) Sensorimotor
adaptation to destabilizing dynamics in weakly electric fish. Current
Biology. link,
DOI:10.1016/j.cub.2024.04.019.
|
|
Biswas, D., Lamperski, A., Yang, Y., Hoffman, K., Guckenheimer, J.,
Fortune, E.S. and N.J. Cowan (2023) Mode switching in organisms for
solving explore-versus-exploit problems. Nature Mach. Intell. link, DOI:10.1038/s42256-023-00745-y.
|
|
Soares, D., Gallman, K., Bichuette, M.E. and E.S. Fortune (2023)
Adaptive shift of active electroreception in weakly electric fish for
troglobitic life. Front. Ecol. Evol. link, DOI:10.3389/fevo.2023.1180506.
|
|
Amendolara, A.B., Sant, B., Rotstein, H.G. and E.S. Fortune (2023)
LSTM-based recurrent neural network provides effective short term flu
forecasting. BMC Public Health, 23:1788 link, PMID:37710241.
|

|
Coleman, M.J., Day, N.F., and E.S. Fortune (2022) Neural mechanisms for
turn-taking in duetting plain-tailed wrens. Front. Neural Circuits,
fn-cir.2022.970434. link, PMID:34074755.
|
|
Coleman, M.J., Day, N.F., Rivera-Parra, P., and E.S. Fortune (2021)
Neurophysiological coordination of duet singing. PNAS,
118(23):e2018188118 link,
PMID:34074755.
|
|
Fortune, E.S., Andanar, N., Madhav, M., Jayakumar, R.P., Cowan, N.J.,
Bichuette, M.E., and D. Soares (2020) Spooky Interaction at a Distance
in Cave and Surface Dwelling Electric Fishes. Front Integr Neurosci,
14:561524. link,
PMID:33192352.
|
|
Gallman, K., Fortune, E.S., Rivera, D., and D. Soares (2020) Differences
in behavior between surface and cave Astyanax mexicanus may be mediated
by changes in catecholamine signaling. J Comp Neurol, 528(16):2639-2653.
link,
PMID:32291742.
|

|
Uyanik, I., Sefati, S., Stamper, S.A., Cho, K-A, Ankarali, M.M.,
Fortune, E.S., and N.J. Cowan (2020) Variability in locomotor dynamics
reveals the critical role of feedback in task control. eLife, e51219. link, PMID:31971509.
|

|
Elie, J.E., Hoffmann, S., Dunning, J.L., Coleman, M.J., Fortune, E.S.,
and J.F. Prather (2019) From Perception to Action: The Role of Auditory
Input in Shaping Vocal Communication and Social Behaviors in Birds.
Brain Behav. Evol., link, PMID:31805560.
|

|
Uyanik, I., Stamper, S.A., Cowan, N.J., and E.S. Fortune (2019) Sensory
Cues Modulate Smooth Pursuit and Active Sensing Movements. Front. Behav.
Neurosci., link,
PMID:31024269.
|
|
Biswas, D., Arend, L.A., Stamper, S.A., Vagvolgyi, B.P., Fortune, E.S.,
and N.J. Cowan. (2018) Closed-Loop Control of Active Sensing Movements
Regulates Sensory Slip. Curr. Biol., 28(24):4029-4036.e4, PMID:30503617.
|

|
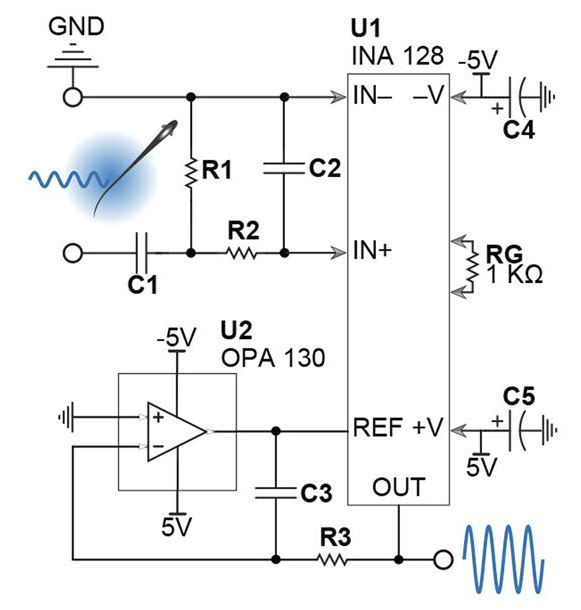
Coleman, M.J. and E.S. Fortune (2018) Quick Guide: Duet Singing in
plain-tailed wrens. Curr. Biol., doi.org/10.1016/j.cub.2018.02.066, pdf.
|
|
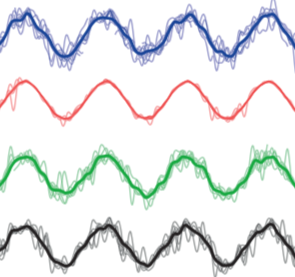
Madhav, M.S., Jayakumar R.P., Demir, A., Stamper S.A., Fortune, E.S.,
and N.J. Cowan (2018) High-resolution behavioral mapping of electrical
fishes in Amazonian habitats. Scientific Reports. link, PMID:29643472.
|
|
Sutton, E.E., Demir, A., Stamper, S.A., Fortune, E.S., and N.J. Cowan
(2016) Dynamic modulation of visual and electrosensory gains for
locomotor control. J R Soc Interface. 13(118).pii:20160057, PMID:27170650.
|
|
Rouse, M.L., Stevenson, T.J., Fortune, E.S., and G.F. Ball (2015)
Reproductive state modulates testosterone-induced singing in adult
female European starlings (Sturnus vulgaris). Horm. Behav.,
72:78-87, PMID:25989596.
|
|
Cowan, N.J., Ankarali, M.M., Dyhr, J.P., Madhav, M.S., Roth, E., Sefati,
S., Stamper, S.A., Fortune, E.S., and T.L. Daniel (2014) Feedback
control as a framework for understanding tradeoffs in biology. Integr.
Comp. Biol., 54:223-237, PMID:23893678.
|
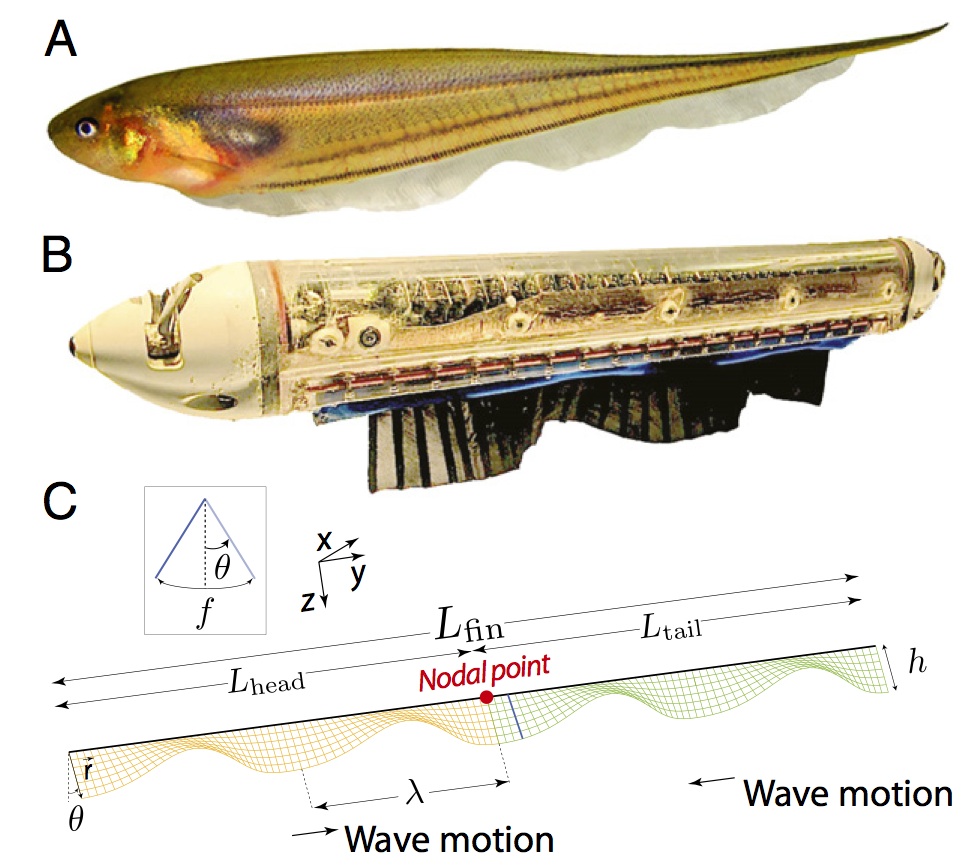
|
Sefati, S., Neveln I.D., Roth E., Mitchell T.R., Snyder J.B., Maciver
M.A., Fortune E.S., and N.J. Cowan (2013) Mutually opposing forces
during locomotion can eliminate the tradeoff between maneuverability and
stability. PNAS, 110:19798-18803, PMID:24191034.
|
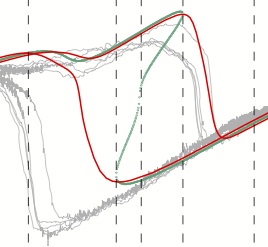
|
Madhav M.S., Stamper, S.A., Fortune, E.S., and N.J. Cowan (2013)
Closed-loop stabilization of the Jamming Avoidance Response reveals its
locally unstable and globally nonlinear dynamics., J. Exp. Biol.,
216:4272-4284, PMID:23997196.
|
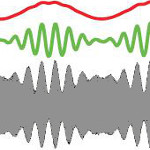
|
Stamper, S.A., Fortune, E.S., and M.J. Chacron (2013) Perception and
coding of envelopes in weakly electric fishes. J. Exp. Biol.,
216:2393-2402, PMID:23761464.
|
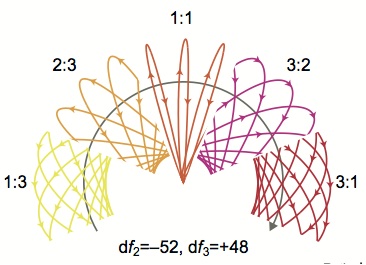
|
Stamper S.A., Madhav M.S., Cowan N.J., and Fortune E.S. (2012) Beyond
the Jamming Avoidance Response: weakly electric fish respond to the
envelope of social electrosensory signals, J. Exp. Biol., 215:4196-4207,
PMID:23136154.
|
|
McGilligray, P., Vonderschen, K., Fortune, E.S., and M.J. Chacron (2012)
Parallel coding of first- and second-order stimulusattributes by
midbrain electrosensory neurons. J. Neurosci., 32:5510-5524, PMID:22514313.
|
|
Stamper, S.A., Roth, E., Cowan, N.J., and E.S. Fortune (2012) Active
sensing via movement shapes spatiotemporal patterns of sensory feedback.
J. Exp. Biol., 215:1567-1574, PMID:22496294.
|
|
Fortune, E.S., Rodriguez, C., Li, D., Ball, G.F., and M.J. Coleman
(2011) Neural mechanisms for the coordination of duet singing in wrens.
Science., 334:666-669, PMID:22052048.
|

|
Roth, E., Zhuang, K., Stamper, S.A., Fortune, E.S., and N.J. Cowan
(2011) Stimulus predictability mediates a switch in locomotor smooth
pursuit performance for Eigenmannia virescens., PMID:21389203.
|
|
Khosravi-Hashemi, N., Fortune, E.S., and M.J. Chacron (2011) Coding
movement direction by burst firing in electrosensory neurons., PMID:21775723.
|
|
Chacron, M.J. and E.S. Fortune (2010) Subthreshold membrane conductances
enhance directional selectivity in vertebrate sensory neurons. J.
Neurophysiol., 4(2):e32, PMID:20445028.
|
|
Stamper, S.A., Carrera-G, E., Tan, E.W., Fugere, V., Krahe, R., and E.S.
Fortune (2010) Species differences in group size and electrosensory
interference in weakly electric fishes: Implications for electrosensory
processing. Behav Brain Res., PMID:19874855.
|
|
Chacron, M.J., Toporikova, N., and E.S. Fortune (2009) Differences in
the time course of short-term depression across receptive fields are
correlated with directional selectivity in electrosensory neurons., J.
Neurophysiol., 102:3270-3279, PMID:19793877.
|
|
Hitschfeld, É.M., Stamper, S.A., Vonderschen, K., Fortune, E.S., and
M.J. Chacron (2009) Effects of restraint and immobilization on
electrosensory behavior of weakly electric fish. ILAR J., 50:361-372, PMID:19949252.
|
|
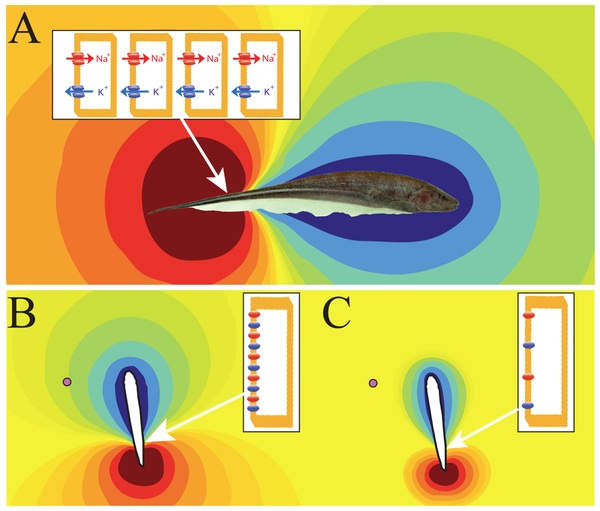
Fortune, E.S. and M.J. Chacron (2009) From molecules to behavior:
organismal-level regulation of ion channel trafficking. PLoS Biol.,
7:e1000211, link,
PMID:19787034.
|

|
Carver, S., Roth, E., Cowan, N.J. and E.S. Fortune (2008) Synaptic
plasticity can produce and enhance direction selectivity. PLoS Comput.
Biol., 4(2):e32, link,
PMID:18282087.
|
|
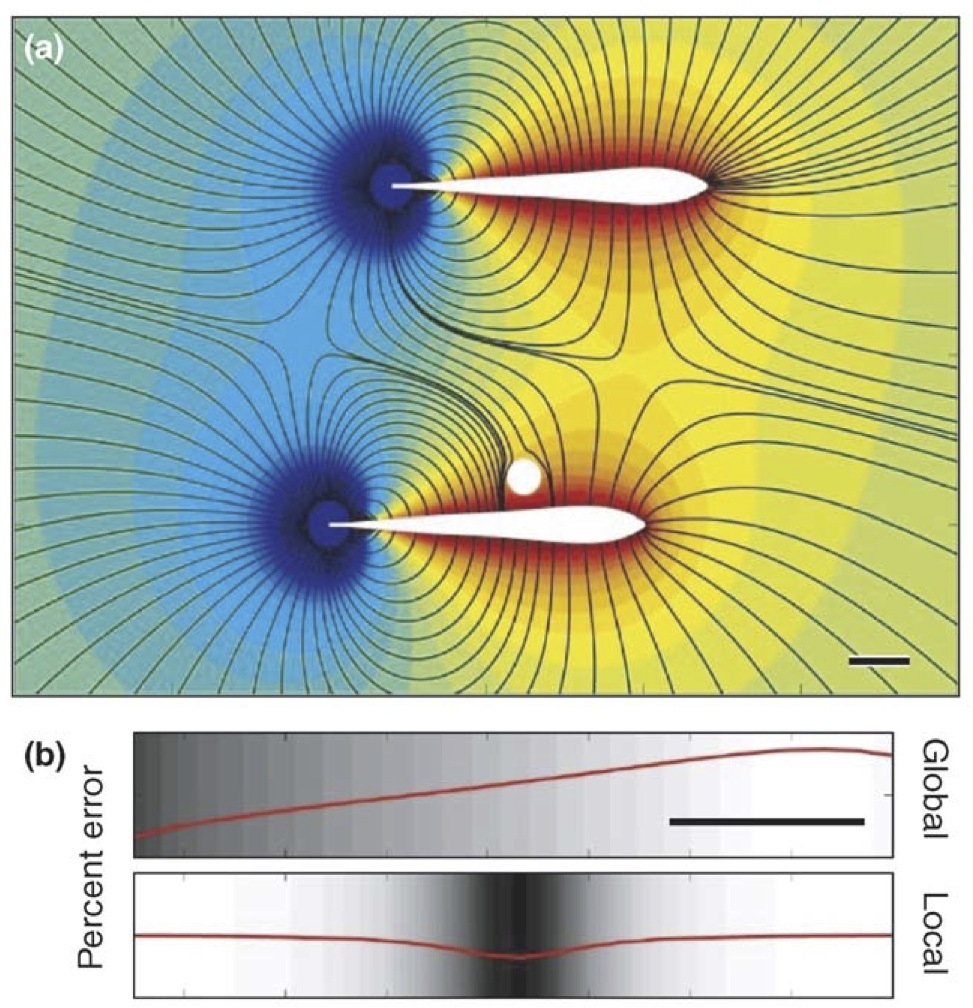
Cowan, N.J. and E.S. Fortune (2007) The critical role of locomotion
mechanics in decoding sensory systems. J. Neurosci, 27:1123-1128, PMID:17267567.
|
|
Fortune, E.S. (2006) The decoding of electrosensory systems. Curr. Opin.
Neurobiol., PMID:16837187.
|
|
Ramcharitar, J.U., Tan, E.W., E.S. Fortune (2006) Global electrosensory
oscillations enhance directional responses of midbrain neurons in
Eigenmannia. J. Neurophys., PMID:16790600.
|
|
Fortune, E.S., Rose, G.J., and M. Kawasaki (2006) Encoding and
processing biologically relevant temporal information in electrosensory
systems. J. Comp. Physiol. A, 192:625-635, PMID:16450118.
|
|
Ramcharitar, J.U., Tan, E.W., and E.S. Fortune (2005)
Effects of global electrosensory signals on motion processing in the
midbrain of Eigenmannia. J. Comp. Physiol. A, 191:865-872, PMID:16001182.
|

|
Tan, E.W., Nizar, J.M., Carrera-G, E., and E.S. Fortune (2005)
Electrosensory interference in naturally occurring aggregates of a
species of weakly electric fish, Eigenmannia virescens. Behav.
Brain Res., 164:83-92, PMID:16099058.
|
|
Fortune, E.S. and G.J. Rose (2003) Voltage-gated Na+ channels
enhance the temporal filtering properties of electrosensory neurons in
the torus. J. Neurophys., 90:924-929, PMID:12750421.
|
|
Fortune, E.S. and G.J. Rose (2002) Roles for short-term synaptic
plasticity in behavior. J Physiol Paris, 96:539-545, PMID:14692501.
|
|
Fortune, E.S. and G.J. Rose (2001) Short-term synaptic plasticity as a
temporal filter. Trends in Neurosciences, 24:381-385, PMID:11410267.
|
|
Rose, G.J. and E.S. Fortune (1999) Frequency-dependent PSP depression
contributes to low-pass temporal filtering in Eigenmannia. J.
Neurosci., 19:7629-7639, PMID:10460268.
|
|
Rose, G.J. and E.S. Fortune (1999) Mechanisms for generating temporal
filters in the electrosensory system. J. Exp. Biol., 202:1281-1289, PMID:10210668.
|
|
Fortune, E.S. and G.J. Rose (1997) Passive and active membrane
properties contribute to the temporal filtering properties of midbrain
neurons, in vivo. J. Neurosci., 17:3815-3825, PMID:9133400.
|
|
Fortune, E.S. and G.J. Rose (1997) Temporal filtering properties of
ampullary electrosensory neurons in the torus semicircularis of
Eigenmannia: evolutionary and computational implications. Brain,
Behav., and Evol., 49:312-323, PMID:9167857.
|
|
Rose, G.J. and E.S. Fortune (1996) New techniques for making whole-cell
recordings from CNS neurons in vivo. Neurosci. Res. 26:89-94, PMID:8895897.
|
|
Fortune, E.S. and D. Margoliash (1995) Parallel pathways and convergence
onto HVc and adjacent neostriatum of adult male zebra finches
(Taeniopygia guttata). J. Comp. Neurol. 360(3):413-441, PMID:8543649.
|
|
Margoliash, D., E.S. Fortune, M. Sutter, C-H. Yu, D. Hardin, and A.Dave
(1994) Distributed representation in the song system of oscines:
evolutionary implications and functional consequences. Brain Behav.
Evol. 44:247-264, PMID:7842284.
|
|
Fortune, E.S. and D. Margoliash (1992) Cytoarchitectonic organization
and morphology of cells of the field L complex in adult male zebra
finches (Taeniopygia guttata). J. Comp. Neurol., 325:388-404, PMID:1447407.
|
|
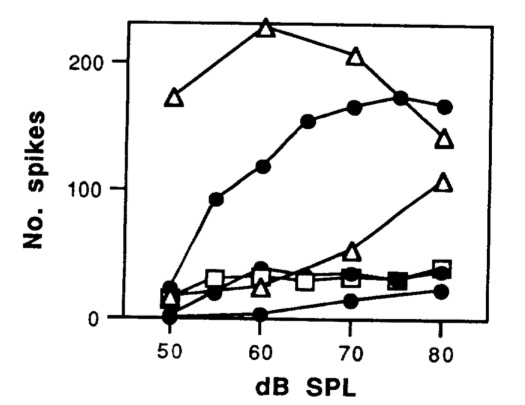
Margoliash, D. and E.S. Fortune (1992) Temporal and harmonic
combination-sensitive neurons in the zebra finch’s HVc. J. Neurosci.,
12:4309-4326, PMID:1432096.
|
Conference Papers
|
Carver, S.G., Fortune, E.S., and N.J. Cowan (2013) State-estimation and
cooperative control with uncertain time. In Proc Amer Control Conf,
Washington, DC, USA, 2013, p. 2990–2995, PDF.
|

|
Sefati, S., Neveln, I., MacIver, M.A., Fortune, E.S., and N.J. Cowan
(2012) Counter-propagating waves enhance maneuverability and stability:
a bio-inspired strategy for robotic ribbon-fin propulsion. In Proc IEEE
Int Conf on Biomed Robot and Biomech, Ro{Mech Eng}, Italy, PDF.
|
Book Chapters

|
Stamper, S.A., Madhav, M.S., Cowan, N.J., and E.S. Fortune (2019) Using
Control Theory to Characterize Active Sensing in Weakly Electric Fishes,
In Springer Handbook of Auditory Research, Volume 70, Electroreception:
Fundamental Insights from Comparative Approaches, 2019, ISBN:
978-3-030-29104-4, StamperUsing2019.
|

|
Metzen, M.G., Fortune, E.S. and M.J. Chacron (2017) Physiology of
Tuberous Electrosensory Systems, In Reference Module in Life Sciences,
Elsevier, 2017, ISBN: 978-0-12-809633-8, MetzenPhysiology2010.
|

|
Fortune, E.S. and N.J. Cowan (2010) Robot Behavior, in The Encyclopedia
of Animal Behavior, eds. Breed and Moore, Elsevier. FortuneRobot2010.
|
|